Plant Physiol. 2024 Apr 12:kiae213.
Fuster-Pons A, Murillo-Sánchez A, Méndez-Vigo B, Marcer A, Pieper B, Torres-Pérez R, Oliveros JC, Tsiantis M, Picó FX, Alonso-Blanco C.
Abstract
Natural variation in trichome pattern (amount and distribution) is prominent among populations of many angiosperms. However, the degree of parallelism in the genetic mechanisms underlying this diversity and its environmental drivers in different species remain unclear. To address these questions, we analyzed the genomic and environmental bases of leaf trichome pattern diversity in Cardamine hirsuta, a relative of Arabidopsis (Arabidopsis thaliana). We characterized 123 wild accessions for their genomic diversity, leaf trichome patterns at different temperatures, and environmental adjustments. Nucleotide diversities and biogeographical distribution models identified two major genetic lineages with distinct demographic and adaptive histories. Additionally, C. hirsuta showed substantial variation in trichome pattern and plasticity to temperature. Trichome amount in C. hirsuta correlated positively with spring precipitation but negatively with temperature, which is opposite to climatic patterns in A. thaliana. Contrastingly, genetic analysis of C. hirsuta glabrous accessions indicated that, like for A. thaliana, glabrousness is caused by null mutations in ChGLABRA1 (ChGL1). Phenotypic genome-wide association studies (GWAS) further identified a ChGL1 haplogroup associated with low trichome density and ChGL1 expression. Therefore, a ChGL1 series of null and partial loss-of-function alleles accounts for the parallel evolution of leaf trichome pattern in C. hirsuta and A. thaliana. Finally, GWAS also detected other candidate genes (e.g. ChETC3, ChCLE17) that might affect trichome pattern. Accordingly, the evolution of this trait in C. hirsuta and A. thaliana shows partially conserved genetic mechanisms but is likely involved in adaptation to different environments.
- Las plantas adaptan su desarrollo y morfología según las condiciones ambientales de su entorno, como la duración del día y la temperatura
- La proteína COP1, además de regular el desarrollo de las plantas en la oscuridad, es responsable de su crecimiento en respuesta a temperaturas ambientes elevadas
- Un nuevo modelo basado en la regulación de COP1 permite predecir cómo el calentamiento global afecta el crecimiento de las plantas según la estación del año y latitud geográfica
Las plantas adaptan su desarrollo y morfología a las condiciones ambientales que las rodean, fundamentalmente, la duración del día y la temperatura ambiente. Estos dos factores afectan de manera directa al rendimiento de los cultivos, de ahí el interés de la comunidad científica en su estudio. Un trabajo de investigadores del Consejo Superior de Investigaciones Científicas (CSIC) presenta un modelo matemático basado en los procesos que modulan a los reguladores de la respuesta a temperatura en plantas, el cuál se ha entrenado con un notable número de datos experimentales.
Esta investigación, publicada en la revista Science Advances, es fruto de la colaboración entre los grupos dirigidos por Salomé Prat y Saúl Ares en el Centro Nacional de Biotecnología perteneciente al CSIC (CNB-CSIC) y Pablo Catalán del Grupo Interdisciplinar de Sistemas Complejos (GISC) de la Universidad Carlos III de Madrid.
Plant Cell Physiol. 2020 Mar 27. pii: pcaa041
Fernández-Santos R, Izquierdo Y, López A, Muñiz L, Martínez M, Cascón T, Hamberg M, Castresana C.
Abstract
Lipid Droplets (LDs) have classically been viewed as seed storage particles, yet they are now emerging as dynamic organelles associated with developmental and stress responses. Nevertheless, their involvement in plant immunity has still been little studied. Here, we found LD accumulation in Arabidopsis thaliana leaves that induced a hypersensitive defense response (HR) after Pseudomonas infection. We established a protocol to reproducibly isolate LDs and to analyze their protein content. The expression of GFP fusion proteins in Nicotiana benthamiana and in transgenic Arabidopsis lines validated the LD localization of glycerol-3-phosphate-acyltransferase 4 (GPAT4) and 8 (GPAT8), required for cutin biosynthesis. Similarly, we showed LD localization of α-dioxygenase1 (α-DOX1) and caleosin3 (CLO3), involved in the synthesis of fatty acid derivatives, and that of phytoalexin deficient 3 (PAD3), which is involved in camalexin synthesis. We found evidence suggesting the existence of different populations of LDs, with varying protein content and distributions. GPAT4 and GPAT8 were associated with LDs inside stomata and surrounding cells of untreated leaves, yet they were mainly confined to LDs in guard cells after bacterial inoculation. By contrast, α-DOX1 and PAD3 were associated with LDs in the epidermal cells of HR-responding leaves, with PAD3 mostly restricted to cells near dead tissue, while CLO3 had a more ubiquitous distribution. As such, the nature of the proteins identified, together with the phenotypic examination of selected mutants, suggest that LDs participate in lipid changes and in the production and transport of defense components affecting the interaction of plants with invading pathogens.
doi: 10.1093/pcp/pcaa041
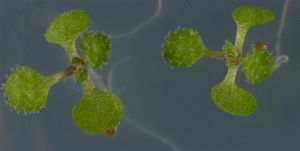
- La regulación de la señalización mediada por ácido jasmónico es clave para mantener el equilibrio entre crecimiento y defensa
- Las proteínas BPM regulan la degradación de los factores de transcripción que activan la respuesta inmune.
La supervivencia de las plantas en la naturaleza requiere su adaptación a diferentes tipos de estrés medioambiental. A diferencia de los animales, las plantas no tienen posibilidad de escapar cuando se enfrentan a una agresión externa como la mordedura de un insecto. Por este motivo, han desarrollado mecanismos de señalización interna y respuesta a través de hormonas como el ácido jasmónico. Sin embargo, la respuesta desencadenada por esta hormona frena el crecimiento de la planta para centrar los recursos en la defensa. Así, un exceso de acción de esta hormona puede afectar al desarrollo vegetal, por lo que su señalización debe estar regulada de una manera muy precisa en tiempo e intensidad.
Ahora, investigadores del Centro Nacional de Biotecnología (CNB) pertenecientes al Consejo Superior de Investigaciones Científicas (CSIC) en colaboración con las universidades de Estrasburgo y Navarra han identificado un nuevo mecanismo molecular que modula los pulsos de activación de la hormona. El trabajo, que ha sido publicado en la revista Proceedings of the National Academy of Sciences (PNAS), da respuesta a cómo se regula la activación de defensas hasta alcanzar un equilibrio entre crecimiento y respuesta a estrés.
PLoS Genet. 2018 Mar 23;14(3):e1007296.
Yang Y, Nicolas M, Zhang J, Yu H, Guo D, Yuan R, Zhang T, Yang J, Cubas P, Qin G.
TIE1 promueve la ramificación de Arabidopsis, dando lugar a plantas más frondosas. Realiza esta función a través de la interacción con BRANCHED1, otra proteína ampliamente estudiada por su papel como modulador de la arquitectura de las plantas. Conocer los procesos que modulan la ramificación en especies de cultivo puede ser útil para optimizar la producción agrícola.
Sánchez-Bermejo E, Castrillo G, Navarro C, del Llano B, Zarco-Fernández S, Martinez-Herrera DJ, Leo-del Puerto Y, Muñoz R, Cámara C, Paz-Ares J, Alonso-Blanco C, Leyva A.
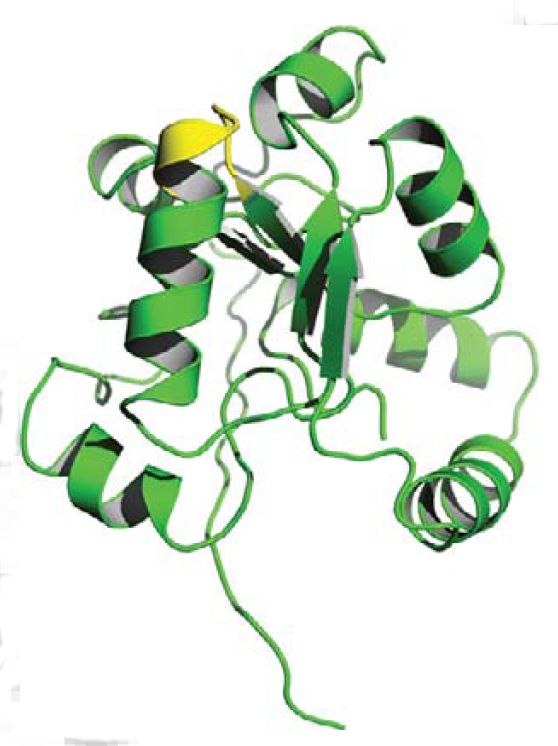 The enormous amount of environmental arsenic was a major factor in determining the biochemistry of incipient life forms early in the Earth’s history. The most abundant chemical
form in the reducing atmosphere was arsenite, which forced organisms to evolve strategies to manage this chemical species. Following the great oxygenation event, arsenite oxidized to arsenate and the action of arsenate reductases became a central survival requirement. The identity of a biologically relevant arsenate reductase in plants nonetheless continues to be debated.
The enormous amount of environmental arsenic was a major factor in determining the biochemistry of incipient life forms early in the Earth’s history. The most abundant chemical
form in the reducing atmosphere was arsenite, which forced organisms to evolve strategies to manage this chemical species. Following the great oxygenation event, arsenite oxidized to arsenate and the action of arsenate reductases became a central survival requirement. The identity of a biologically relevant arsenate reductase in plants nonetheless continues to be debated.
Here we identify a quantitative trait locus that encodes a novel arsenate reductase critical for arsenic tolerance in plants. Functional analyses indicate that several non-additive polymorphisms affect protein structure and account for the natural variation in arsenate reductase activity in Arabidopsis thaliana accessions. This study shows that arsenate reductases are an essential component for natural plant variation in As(V) tolerance.
Manzano-Piedras E, Marcer A, Alonso-Blanco C, Picó FX.
The role that different life-history traits may have in the process of adaptation caused by divergent selection can be assessed by using extensive collections of geographically-explicit populations. This is because adaptive phenotypic variation shifts gradually across space as a result of the geographic patterns of variation in environmental selective pressures. Hence, large-scale experiments are needed to identify relevant adaptive life-history traits as well as their relationships with putative selective agents.
We conducted a field experiment with 279 geo-referenced accessions of the annual plant Arabidopsis thaliana collected across a native region of its distribution range, the Iberian Peninsula. We quantified variation in life-history traits throughout the entire life cycle. We built a geographic information system to generate an environmental data set encompassing climate, vegetation and soil data. We analysed the spatial autocorrelation patterns of environmental variables and life-history traits, as well as the relationship between environmental and phenotypic data. Almost all environmental variables were significantly spatially autocorrelated. By contrast, only two life-history traits, seed weight and flowering time, exhibited significant spatial autocorrelation. Flowering time, and to a lower extent seed weight, were the life-history traits with the highest significant correlation coefficients with environmental factors, in particular with annual mean temperature. In general, individual fitness was higher for accessions with more vigorous seed germination, higher recruitment and later flowering times. Variation in flowering time mediated by temperature appears to be the main life-history trait by which A. thaliana adjusts its life history to the varying Iberian environmental conditions.
The use of extensive geographically-explicit data sets obtained from field experiments represents a powerful approach to unravel adaptive patterns of variation. In a context of current global warming, geographically-explicit approaches, evaluating the match between organisms and the environments where they live, may contribute to better assess and predict the consequences of global warming.
BMC Plant Biol. 2014; 14(1): 17.
Brennan AC, Méndez-Vigo B, Haddioui A, Martínez-Zapater JM, Picó FX, Alonso-Blanco C.
 Deciphering the genetic structure of Arabidopsis thaliana diversity across its geographic range provides the bases for elucidating the demographic history of this model plant. Despite the unique A. thaliana genomic resources currently available, its history in North Africa, the extreme southern limit in the biodiversity hotspot of the Mediterranean Basin, remains virtually unknown.
Deciphering the genetic structure of Arabidopsis thaliana diversity across its geographic range provides the bases for elucidating the demographic history of this model plant. Despite the unique A. thaliana genomic resources currently available, its history in North Africa, the extreme southern limit in the biodiversity hotspot of the Mediterranean Basin, remains virtually unknown.
To approach A. thaliana evolutionary history in North Africa, we have analysed the genetic diversity and structure of 151 individuals collected from 20 populations distributed across Morocco. Genotyping of 249 genome-wide SNPs indicated that Morocco contains substantially lower diversity than most analyzed world regions. However, IBD, STRUCTURE and PCA clustering analyses showed that genetic variation is strongly geographically structured. We also determined the genetic relationships between Morocco and the closest European region, the Iberian Peninsula, by analyses of 201 populations from both regions genotyped with the same SNPs. These analyses detected four genetic groups, but all Moroccan accessions belonged to a common Iberian/Moroccan cluster that appeared highly differentiated from the remaining groups. Thus, we identified a genetic lineage with an isolated demographic history in the south-western Mediterranean region. The existence of this lineage was further supported by the study of several flowering genes and traits, which also found Moroccan accessions similar to the same Iberian group. Nevertheless, genetic diversity for neutral SNPs and flowering genes was higher in Moroccan than in Iberian populations of this lineage. Furthermore, we analyzed the genetic relationships between Morocco and other world regions by joint analyses of a worldwide collection of 337 accessions, which detected an additional weak relationship between North Africa and Asia.
The patterns of genetic diversity and structure of A. thaliana in Morocco show that North Africa is part of the species native range and support the occurrence of a glacial refugium in the Atlas Mountains. In addition, the identification of a genetic lineage specific of Morocco and the Iberian Peninsula indicates that the Strait of Gibraltar has been an A. thaliana migration route between Europe and Africa. Finally, the genetic relationship between Morocco and Asia suggests another migration route connecting north-western Africa and Asia
Journal of Experimental Botany
Méndez-Vigo B, Andrés MT, Ramiro M, Martínez-Zapater JM & Alonso-Blanco C.
Vegetative growth and flowering initiation are two crucial developmental processes in the life cycle of annual plants that are closely associated. The timing of both processes affects several presumed adaptive traits, such as flowering time (FT), total leaf number (TLN), or the rate of leaf production (RLP). However, the interactions among these complex processes and traits, and their mechanistic bases, remain largely unknown. To determine the genetic relationships between them, the natural genetic variation between A. thaliana accessions Fei-0 and Ler has been studied using a new population of 222 LerxFei-0 recombinant inbred lines. Temporal analysis of the parental development under a short day photoperiod distinguishes two vegetative phases differing in their RLP. QTL mapping of RLP in consecutive time intervals of vegetative development indicates that Ler/Fei-0 variation is caused by 10 loci whose small to moderate effects mainly display two different temporal patterns. Further comparative QTL analyses show that most of the genomic regions affecting FT or TLN also alter RLP. In addition, the partially independent genetic bases observed for FT and TLN appear determined by several genomic regions with two different patterns of phenotypic effects: regions with a larger effect on FT than TLN, and vice versa. The distinct temporal and pleiotropic patterns of QTL effects suggest that natural variation for flowering time is caused by different genetic mechanisms involved in vegetative and/or reproductive phase changes, most of them interacting with the control of leaf production rate. Thus, natural selection might contribute to maintain this genetic variation due to its phenotypic effects not only on the timing of flowering initiation but also on the rate of vegetative growth.POLÍTICA DE COOKIES
Una cookie es un archivo de texto que se almacena en el ordenador o dispositivo móvil mediante un servidor Web y tan solo ese servidor será capaz de recuperar o leer el contenido de la cookie y permiten al Sitio Web recordar preferencias de navegación y navegar de manera eficiente. Las cookies hacen la interacción entre el usuario y el sitio Web más rápida y fácil.
Información general
Está página Web utiliza cookies. Las cookies son pequeños archivos de texto generados por las páginas web que usted visita, las cuales contienen los datos de sesión que pueden ser de utilidad posteriormente en la página web. De esta forma esta Web recuerda información sobre su visita, lo que puede facilitar su próxima visita y hacer que el sitio Web le resulte más útil.
¿Cómo funcionan las cookies?
Las cookies sólo pueden almacenar texto, por lo general siempre es anónimo y cifrado. No se almacenarán información personal alguna en una cookie, ni pueden asociarse a persona identificada o identificable.
Los datos permiten que esta Web pueda mantener su información entre las páginas, y también para analizar la forma de interactuar con el sitio Web. Las cookies son seguras ya que sólo pueden almacenar la información que se puso en su lugar por el navegador, lo que es información que el usuario ha introducido en el navegador o la que se incluye en la solicitud de página. No puede ejecutar el código y no se puede utilizar para acceder a su ordenador. Si una página web cifra la información de la cookie, sólo la página web puede leer la información.
¿Qué tipos de cookies utilizamos?
Las cookies que utiliza esta página Web se pueden distinguir según los siguientes criterios:
1. Tipos de cookies según la entidad que las gestiona:
Según quien sea la entidad que gestione el equipo o dominio desde donde se envían las cookies y trate los datos que se obtengan, podemos distinguir:
- Cookies propias: son aquellas que se envían al equipo terminal del usuario desde un equipo o dominio gestionado por el propio editor y desde el que se presta el servicio solicitado por el usuario.
- Cookies de terceros: son aquellas que se envían al equipo terminal del usuario desde un equipo o dominio que no es gestionado por el editor, sino por otra entidad que trata los datos obtenidos través de las cookies.
En el caso de que las cookies sean instaladas desde un equipo o dominio gestionado por el propio editor pero la información que se recoja mediante éstas sea gestionada por un tercero, no pueden ser consideradas como cookies propias.
2. Tipos de cookies según el plazo de tiempo que permanecen activadas:
Según el plazo de tiempo que permanecen activadas en el equipo terminal podemos distinguir:
- Cookies de sesión: son un tipo de cookies diseñadas para recabar y almacenar datos mientras el usuario accede a una página web. Se suelen emplear para almacenar información que solo interesa conservar para la prestación del servicio solicitado por el usuario en una sola ocasión (p.e. una lista de productos adquiridos).
- Cookies persistentes: son un tipo de cookies en el que los datos siguen almacenados en el terminal y pueden ser accedidos y tratados durante un periodo definido por el responsable de la cookie, y que puede ir de unos minutos a varios años.
3. Tipos de cookies según su finalidad:
Según la finalidad para la que se traten los datos obtenidos a través de las cookies, podemos distinguir entre:
- Cookies técnicas: son aquellas que permiten al usuario la navegación a través de una página web, plataforma o aplicación y la utilización de las diferentes opciones o servicios que en ella existan como, por ejemplo, controlar el tráfico y la comunicación de datos, identificar la sesión, acceder a partes de acceso restringido, recordar los elementos que integran un pedido, realizar el proceso de compra de un pedido, realizar la solicitud de inscripción o participación en un evento, utilizar elementos de seguridad durante la navegación, almacenar contenidos para la difusión de vídeos o sonido o compartir contenidos a través de redes sociales.
- Cookies de personalización: son aquellas que permiten al usuario acceder al servicio con algunas características de carácter general predefinidas en función de una serie de criterios en el terminal del usuario como por ejemplo serian el idioma, el tipo de navegador a través del cual accede al servicio, la configuración regional desde donde accede al servicio, etc.
- Cookies de análisis: son aquellas que permiten al responsable de las mismas, el seguimiento y análisis del comportamiento de los usuarios de los sitios web a los que están vinculadas. La información recogida mediante este tipo de cookies se utiliza en la medición de la actividad de los sitios web, aplicación o plataforma y para la elaboración de perfiles de navegación de los usuarios de dichos sitios, aplicaciones y plataformas, con el fin de introducir mejoras en función del análisis de los datos de uso que hacen los usuarios del servicio.
Herramienta de gestión de las cookies
Está página Web utiliza Google Analytics.
Google Analytics es una herramienta gratuita de análisis web de Google que principalmente permite que los propietarios de sitios web conozcan cómo interactúan los usuarios con su sitio web. Asimismo, habilita cookies en el dominio del sitio en el que te encuentras y utiliza un conjunto de cookies denominadas "__utma" y "__utmz" para recopilar información de forma anónima y elaborar informes de tendencias de sitios web sin identificar a usuarios individuales.
Para realizar las estadísticas de uso de esta Web utilizamos las cookies con la finalidad de conocer el nivel de recurrencia de nuestros visitantes y los contenidos que resultan más interesantes. De esta manera podemos concentrar nuestros esfuerzos en mejorar las áreas más visitadas y hacer que el usuario encuentre más fácilmente lo que busca. En esta Web puede utilizarse la información de su visita para realizar evaluaciones y cálculos estadísticos sobre datos anónimos, así como para garantizar la continuidad del servicio o para realizar mejoras en sus sitios Web. Para más detalles, consulte en el siguiente enlace la política de privacidad [http://www.google.com/intl/es/policies/privacy/]
Cómo gestionar las cookies en su equipo: la desactivación y eliminación de las cookies
Todos los navegadores de Internet le permiten limitar el comportamiento de una cookie o desactivar las cookies dentro de la configuración o las opciones del navegador. Los pasos para hacerlo son diferentes para cada navegador, se pueden encontrar instrucciones en el menú de ayuda de su navegador.
Si no acepta el uso de las cookies, ya que es posible gracias a los menús de preferencias o ajustes de su navegador, rechazarlas, este sitio Web seguirá funcionando adecuadamente sin el uso de las mismas.
Puede usted permitir, bloquear o eliminar las cookies instaladas en su equipo mediante la configuración de las opciones del navegador instalado en su ordenador:
- Para más información sobre Internet Explorer pulse aquí.
- Para más información sobre Chrome pulse aquí.
- Para más información sobre Safari pulse aquí.
- Para más información sobre Firefox pulse aquí.
A través de su navegador, usted también puede ver las cookies que están en su ordenador, y borrarlas según crea conveniente. Las cookies son archivos de texto, los puede abrir y leer el contenido. Los datos dentro de ellos casi siempre están cifrados con una clave numérica que corresponde a una sesión en Internet por lo que muchas veces no tienen sentido más allá que la página web que los escribió.
Consentimiento informado
La utilización de la presente página Web por su parte, implica que Vd. presta su consentimiento expreso e inequívoco a la utilización de cookies, en los términos y condiciones previstos en esta Política de Cookies, sin perjuicio de las medidas de desactivación y eliminación de las cookies que Vd. pueda adoptar, y que se mencionan en el apartado anterior.



